1. 서 론
국내 전통한지의 주요 원료로 사용되는 닥나무는 비목재 섬유인 닥나무 인피섬유가 사용되며, APG II 분류체계에 의하면 닥나무속(Broussonetia)은 장미군(Rosids), 장미목(Rosales), 뽕나무과(Moraceae)에 속하는 낙엽 활엽 관목으로 알려져 있다.1) 닥나무속의 분자계통학적 연구에 의하면(Plants of the world online) 교잡종 1건을 포함해 총 5종의 닥나무로 분류하고 있으며 다양한 동의어를 포함하고 있다.2) 한국의 닥나무속 연구는 1911년 Nakai가 한반도에 B. papyrifera vent.가 분포한다고 언급함으로써 시작되었다.3) 그 이후 1922년 Mori가 닥나무속에 B. kazinoki Siebold과 B. papyrifera vent.로 2종을 기재하였다.4) 1969년에 닥나무류의 수목학적 특징을 통해 hybrid(B. kazinoki Siebold × B. papyrifera vent.)된 닥나무의 존재를 제기하였음에도 불구하고5) 학술적으로 인정되지 않았고 1980년까지도 2종으로 기록되어 왔다. 2009년에는 가거애기닥나무(B. kazinoki for. Koreana M. Kim)의 새로운 등장과 함께 학명의 변경을 제안하였으며 닥나무속에 대한 다양한 의견들이 제안되고 있는 실정이다. Ohba는 4가지 종을 제안하고 있으며6) 국립수목원에서 지원하는 국가생물종지식정보시스템에서는 비합법명인 꾸지닥나무를 포함하여 4개 종의 닥나무속으로 검색되고 있다(Table 1).7)
Table 1.
Classification of Genus Broussonetia published academically
Plants of the
world online (2022) | Synonyms |
|---|
| B. × kazinoki Siebold | B. × hanjiana M.Kim | B. × kazinoki var.
humilis Uyeki | B. × kazinoki
f. humilis (Uyeki) M.Kim | B. ×
sieboldii Blume | | | |
B.
papyrifera (L.)
L'Hér. ex Vent. | B. billardii Carruth. | B. cordata K.Koch | B. cordata Blume | B. cucullata Steud. | B. dissecta
Bureau | B. elegans
K.Koch | ⋯ |
| B. monoica Hance | B.×kazinoki
f. koreana M.Kim | B. × kazinoki var.
ruyangensis P.H.Liang &
X.W.Wei | B. rupicola
F.T.Wang &
Tang | | | | |
| B. harmandii Gagnep. | | | | | | | |
| B. kaempferi Siebold | Morus
kaempferi (Siebold) Ser. | Papyrius
kaempferi (Siebold)
Kuntze | Papyrius
karinokii Kuntze | | | | |
Plants of the world
online (2022) | Korea national arboretum
(2022) | Ohba
(2014) | Yun & Kim
(2009) | Lee (1980) | Mori (1922) | Nakai (1911) |
|---|
| B. × kazinoki Siebold | B. ×kazinoki M.Kim /
닥나무 | B. kazinoki Siebold | B. kazinoki Siebold /
애기닥나무 | B. kazinoki Sieb. /
닥나무 | B. kazinoki Siebold | |
B. papyrifera (L.)
L'Hér. ex Vent. | B. papyrifera (L.) L'Hér. ex Vent. /
꾸지나무 | B. papyrifera (L.) Vent. | B. papyrifera (L.) L'Hér.
ex Vent. / 꾸지나무 | B. papyrifera (L.)
Vent. / 꾸지나무 | B. papyrifera vent. | B. papyrifera vent. |
| B. monoica Hance | | B. monoica Hance | | | | |
| B. harmandii Gagnep. | | B. kaempferi Siebold | | | | |
| B. kaempferi Siebold | | | | | | |
| B. kazinoki ×
B.
papyrifera /
꾸지닥나무 (비합법명) | | B. × hanjiana M.Kim /
닥나무 | | | |
| B.× kazinoki Siebold
ex Siebold & Zucc. /
애기닥나무 | | | | | |
| | | B. × kazinoki for.
Koreana M. Kim /
가거애기닥나무 | | | |
특히 국내 연구동향으로는 잎의 성상이나 암꽃의 포 길이, 엽신의 폭과 같은 외부 형태학적 특징을 파악한 연구가 대부분이다. 닥나무(B. kazinoki Siebold)와 꾸지나무(B. papyrifera(L.) L’Hér. ex Vent.)의 수목학적 특징이 유사하여 오동정의 사례가 발생하기도 하며8) 닥나무를 잎의 성상에 따라 진저(眞楮), 적저(赤楮), 요저(要楮), 흑저(黑楮) 등으로 구분하기도 하며 국내 한지업계에서는 통상 참닥과 부닥으로 구분하고 있다고 보고되고 있으며,9) 현장에서는 이보다 더 다양한 명칭들로 불리어지고 있다.10,11) 게다가 전통한지에 사용되는 닥나무는 1년 미만생으로 수목학에서 보는 가장 주요한 특징인 꽃과 열매를 확인할 수 없는 한계가 있다.
한지는 주로 닥나무 인피섬유를 가지고 우리나라 고유의 방식으로 만들어진 종이를 일컫는데 고문헌의 기록에서 한지는 사용된 재료, 제조방법, 생산지 등에 따라 다양하게 불리기도 했다. 품질에 따라 공상지(供上紙), 별백지(別白紙), 상백지(常白紙) 등으로 나뉘는데 색상, 양질을 놓고 구분하였다. 종이의 물리적인 강도에는 종이의 지합, 인장 및 인열강도 등이 있으며 이는 섬유장, 폭 등과 같은 섬유의 형태학적 특성이 큰 영향을 미친다. 12-14) 또한 한지를 제작하는 공정변수에도 의존되지만 기본적으로 수종에 따른 섬유의 특성에 따라 현저하게 다를 수 있다고 보고되고 있다.15)
따라서 본 연구는 전통한지 제조에 사용되고 있는 닥나무속 식물들의 종간 유연관계를 확인하여 분류학적 위치의 모호함을 해결하고자 하였다. 나아가 닥나무속의 종간 계통분류학적 유연관계에 따른 닥나무의 섬유의 길이, 폭, 루멘폭의 특징과 비교를 하고자 함에 그 목적이 있다.
2. 재료 및 방법
2.1 공시재료
2.1.1 실험재료
본 연구에 사용된 재료는 2019년 3월부터 2019년 11월에 걸쳐 전국 권역별 4개도에서 지역별로 23개체의 닥나무의 잎과 인피섬유를 직접 생체로 채집하였다. 채집된 닥나무의 채집지역은 Table 2와 같다.
Table 2.
Materials of paper-mulberry used in this study
| Number | CODE | Common name | Locality |
|---|
| 1 | B01 | Chamdak | Gyeongbuk | Mungyeong |
| 2 | B02 | Chamdak | Andong |
| 3 | B03 | Meogudak |
| 4 | B05 | Chamdak | Uiseong |
| 5 | B06 | Gaedak | Uiseong |
| 6 | B07 | Chamdak | Jeonbuk | Jangsu |
| 7 | B08 | Chamdak |
| 8 | B09 | Chamdak |
| 9 | B10 | Chamdak | Imsil |
| 10 | B11 | Jireum |
| 11 | B12 | Meogudak |
| 12 | B13 | Doksae |
| 13 | B14 | Hayan |
| 14 | B15 | Munui |
| 15 | B16 | Ttalgi |
| 16 | B17 | Chamdak | Waemang |
| 17 | B18 | Chamdak | Wanju |
| 18 | B19 | Meogudak | Wanju |
| 19 | B20 | Chamdak | Gyeongbuk | Cheongsong |
| 20 | B21 | Chamdak |
| 21 | B22 | Chamdak |
| 22 | B23 | Chamdak | Gyeonggi | Gapyeong |
| 23 | B24 | Chamdak | Chungbuk | Goesan |
2.2 실험방법
2.2.1 DNA 추출 및 Amplification
재배지에서 채취한 닥나무의 생체 잎을 실험실로 운반하여 닥나무의 잎을 채취한 후 -70℃ 급속냉동고에서 보관하였다. 생체시료를 액체질소와 함께 분쇄한 후 allen et al.(2006)의 CTAB방법을 사용하여 DNA를 추출하였다.16) 추출된 DNA는 1% agarose gel을 이용하여 전기영동을 통해 DNA의 유무를 확인하고 NANO drop으로 DNA 순도(O/D)와 농도를 측정하였다. 식물 종의 유연관계와 진화경향성 및 유전적 변이양상을 파악하기 위해서 ITS 지역, DNA barcoding marker 중 coding region인 matK, ndhF와 non-coding 지역인 trnL-F 지역을 선정하였다.
PCR 반응용액의 조성은 DNA 20-50 ng, 5×DiastarTMTaq DNA 완충액 2.5 μL, 10 mM의 dNTPs 0.5 μL, 10 pmol의 프라이머를 각각 1 μL, 2.5 unit의 DiastarTM Taq DNA Polymerase 0.13 μL(SolGent Co., Korea)를 첨가 후 마지막으로 전체 부피가 25 μL가 되도록 하였다. DNA 증폭을 위한 PCR 반응은 다음과 같다. 조제된 반응용액은 95℃에서 2분 동안 initial denaturation하고, 이후 95℃에서 20초, annealing 56℃ 40초, extension 72℃ 1분으로 구성된 반응을 30-35회 반복 후 최종적으로 72℃에서 5분 간 extension하여 수행하였다. PCR 반응용액은 PCR purification system kit(solgent co., Korea)로 정제하였다. 또한 시퀀싱 분석에서 나타나는 문제를 최소화하고 여러 타입으로 나타나는 염기서열을 분리하고자 각 시료 당 콜로니 8개를 채취하여 시퀀싱 분석을 실시하였다.
2.2.2 염기서열 정렬 및 계통분석
확보한 염기서열들을 Geneious R11 프로그램17)을 사용하여 각 샘플에 대한 염기서열 조합(assembly)하며, Clustal W 프로그램을 사용하여 염기서열을 정렬(Alignment)하였다. 염기서열 간의 유전적거리(Kimura-2-parameter방법)18) 계산과 RAxML v 8.0.0 프로그램19)을 통해 Maximum likelihood(ML)를 분석하였다. 분계도의 신뢰성을 높이기 위해 Best model을 확정하고 bootstrap value를 이용하여 무작위로 500회 반복하였다.
2.2.3 형태학적 관찰
섬유장, 섬유폭을 측정하기 위하여 닥나무 인피섬유를 8 cm로 시료를 절단하였다. 시료를 해리(defibration)하기 위하여 30% nitric acid(60%, Kanto) 100 mL와 potassium chlorate(99%, Sigma Aldrich) 5 g을 혼합하여 Schulze 용액을 제조하였다. 시료를 Schulze 용액에 넣고 5시간 동안 상온에서 방치한 후 증류수로 충분히 수세하고 증류수와 함께 시료를 흔들어 해리 시킨다. 해리된 섬유를 1% safranin 용액으로 염색한 후 프레파라트를 제작하여 광학현미경(Nikon, Eclipse Ni, Japan)으로 관찰하였다. 섬유장, 섬유폭은 각각 25회, 루멘폭은 50회씩 측정하였다.
펄프제지특성을 나타내는 runkel ratio(RR), slenderness ratio(SR), flexibility coefficient(FC), rigidity coefficient(RC) 값은 [1]-[4]수식에 의거하였다.20-22)
3. 결과 및 고찰
3.1 분자계통학적 분석
NCBI 데이터베이스 내 닥나무속 및 근연 분류군의 염기서열 자료를 활용하여 수집한 생체시료 23개체에 대한 ITS, trnL-F, matK, ndhF 지역의 염기서열 비교분석 및 계통학적 분석의 결과는 Fig. 1, Table 3과 같다.
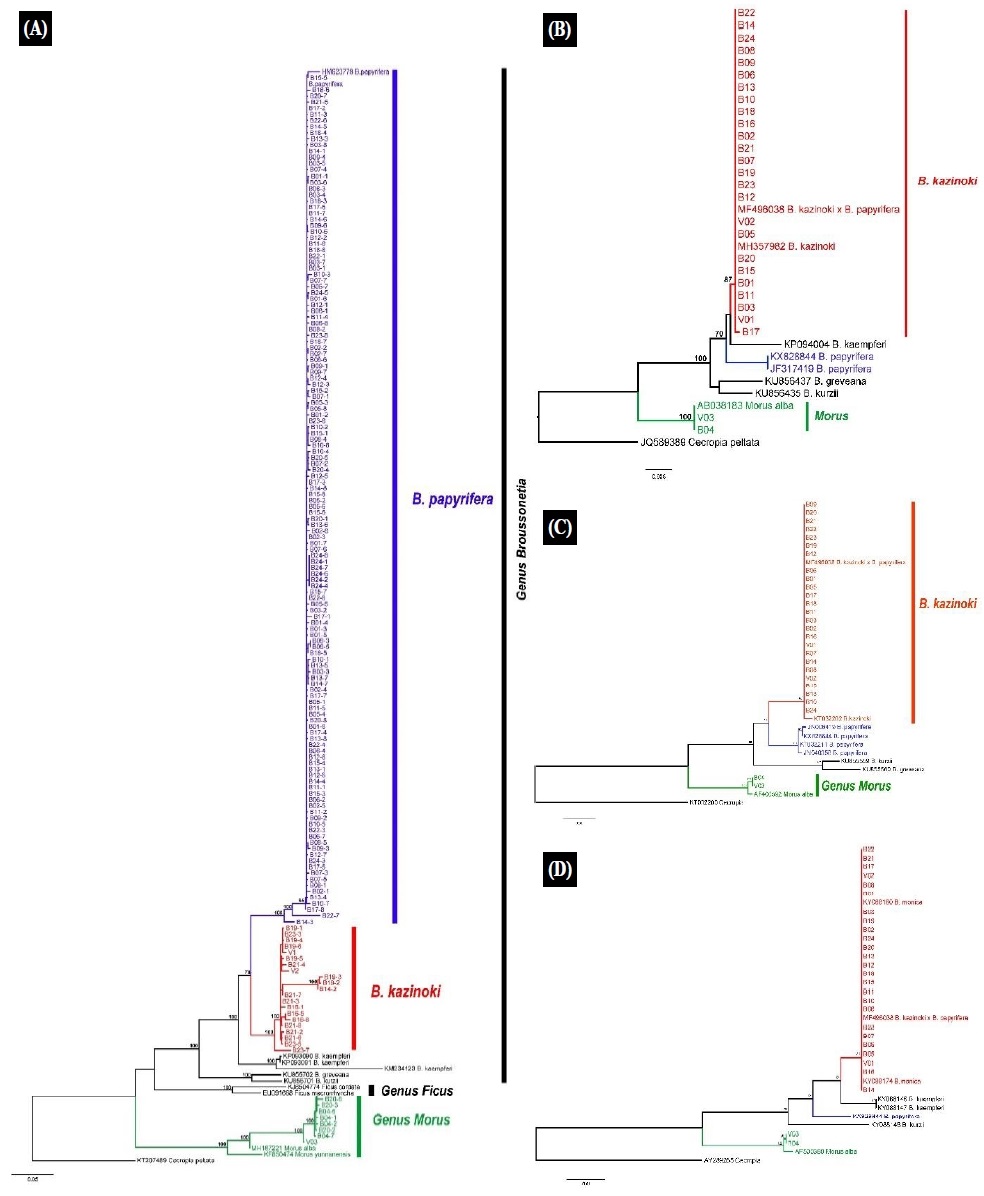
Fig. 1.
Phylogenetic relationships of genus Broussonetia based on the ML tree molecular dataset. Number at the nodes indicate bootstrap values; (A): ITS, (B): Mat K, (C): trnL-F, (D): ndhF.
Table 3.
Results of phylogenetic relationships paper-mulberry based on the combined (ITS, matK, trnL-F, ndhF), Hybrid*; B. kazinoki × B. papyrifera; 1) B14-2 sample formed a boundary line with B. kazinoki; 2) B20-2, 3, 6 samples formed a boundary line with M.alba; 3) B21-5 sample formed a boundary line with B. papyrifera
| Abbreviation | Branch of a tree formed | Results |
|---|
| ITS | trnL-F, matK, ndhF |
|---|
| B01 | B. papyrifera | B. kazinoki | Hybrid* |
| B02 | B. papyrifera | B. kazinoki | Hybrid* |
| B03 | B. papyrifera | B. kazinoki | Hybrid* |
| B05 | B. papyrifera | B. kazinoki | Hybrid* |
| B06 | B. papyrifera | B. kazinoki | Hybrid* |
| B07 | B. papyrifera | B. kazinoki | Hybrid* |
| B08 | B. papyrifera | B. kazinoki | Hybrid* |
| B09 | B. papyrifera | B. kazinoki | Hybrid* |
| B10 | B. papyrifera | B. kazinoki | Hybrid* |
| B11 | B. papyrifera | B. kazinoki | Hybrid* |
| B12 | B. papyrifera | B. kazinoki | Hybrid* |
| B13 | B. papyrifera | B. kazinoki | Hybrid* |
| B14 | B. papyrifera1) | B. kazinoki | Hybrid* |
| B15 | B. papyrifera | B. kazinoki | Hybrid* |
| B16 | B. kazinoki | B. kazinoki | B. kazinoki |
| B17 | B. papyrifera | B. kazinoki | Hybrid* |
| B18 | B. papyrifera | B. kazinoki | Hybrid* |
| B19 | B. kazinoki | B. kazinoki | B. kazinoki |
| B20 | B. papyrifera × M. alba2) | B. kazinoki | Hybrid* |
| B21 | B. papyrifera3) | B. kazinoki | Hybrid* |
| B22 | B. papyrifera | B. kazinoki | Hybrid* |
| B23 | B. papyrifera | B. kazinoki | Hybrid* |
| B24 | B. papyrifera | B. kazinoki | Hybrid* |
ITS 분석 결과 B14-2, B16, B19, B21-2, 3, 4, 6, 7, B23-3, 5, 7이 BS(Bootstrap)값 100%로 닥나무(B. kazinoki)와 분계조를 형성하였다. 그리고 B20-2,3,4 시료가 BS값 100%로 뽕나무(M. alba)와 분계조를 형성하였다. 그 외 B14-1, 3, 4, 5, 6, 7, 8과 B21-5 시료들은 꾸지나무(B. papyrifera)와 분계조를 형성하였다.
trnL-F, matK, ndhF 지역은 모든 시료가 BS값 100%로 닥나무(B. kazinoki)와 분계조를 형성하며 모두 동일한 결과를 나타냈다.
시료 23개를 대상으로 분자마커 4개(ITS, trnL-F, matK, ndhF)를 사용하여 염기서열 비교분석 및 계통학적 분석결과를 종합해 보면, 전체 23개 시료 중 B16과 B19를 제외한 나머지 시료는 엽록체 내 3개지역 trnL-F, matK, ndhF의 분석결과 모두 기존의 닥나무와 동일하거나 유사한 것으로 나타났으며, 핵 내 1개 지역(ITS)의 분석결과에서는 닥나무, 꾸지나무, 뽕나무의 염기서열과 동일하거나 유사한 것으로 나타났다. 이로써 21개체의 hybrid 시료는 닥나무와 꾸지나무의 교잡종(Hybrid = B. kazinoki × B. papyrifera)으로 판단된다. 엽록체 유전체의 경우, 모계유전(Maternal inheritance)을 통해서 유전정보가 전달됨으로 잡종화에서 모계를 찾는데 매우 유용한 유전정보이다. 엽록체 지역의 세 개의 마커에서 시료들은 닥나무와 매우 가까운 것으로 나타나 잡종으로 판단되는 시료의 모계종은 닥나무로 판단된다. B16과 B19는 B. kazinoki로 확인되었다.
3.2 형태학적 분석
23개체 중 22개체 닥나무 인피섬유를 대상으로 섬유장, 섬유폭, 루멘폭 등을 측정한 값은 Table 4와 같다. B19 시료의 경우 관상용으로 직접적인 시료채취가 어려웠기 때문에 형태학적 분석에서 제외되었다.
Table 4.
Analysis of variance for fiber morphology in the bast fibers according to Phylogenetic relationships in the paper-mulberry
| Abbreviation | Classification of
Genus Broussonetia | Length
(mm) | Diameter
(μm) | Lumen width
(μm)
| Cell wall
thickness (μm) |
|---|
| B01 | Hybrid* | 9.01 ± 0.47 | 19.56 ± 0.53 | 6.14 ± 0.64 | 6.71 |
| B02 | Hybrid* | 7.99 ± 0.37 | 18.54 ± 0.42 | 6.46 ± 0.64 | 6.04 |
| B03 | Hybrid* | 8.43 ± 0.39 | 17.34 ± 0.40 | 6.54 ± 0.68 | 5.40 |
| B05 | Hybrid* | 8.54 ± 0.51 | 19.84 ± 0.63 | 7.17 ± 0.75 | 6.34 |
| B06 | Hybrid* | 10.37 ± 0.50 | 21.14 ± 0.75 | 7.92 ± 0.88 | 6.61 |
| B07 | Hybrid* | 9.00 ± 0.32 | 18.85 ± 0.58 | 6.38 ± 0.70 | 6.24 |
| B08 | Hybrid* | 9.49 ± 0.36 | 19.32 ± 0.62 | 6.80 ± 0.74 | 6.26 |
| B09 | Hybrid* | 9.33 ± 0.45 | 19.02 ± 0.52 | 9.85 ± 1.12 | 4.59 |
| B10 | Hybrid* | 9.30 ± 0.29 | 22.22 ± 0.20 | 6.91 ± 0.73 | 7.66 |
| B11 | Hybrid* | 8.37 ± 0.39 | 22.29 ± 0.28 | 7.13 ± 0.80 | 7.58 |
| B12 | Hybrid* | 10.46 ± 0.36 | 22.94 ± 0.30 | 7.30 ± 0.73 | 7.82 |
| B13 | Hybrid* | 9.35 ± 0.46 | 23.64 ± 0.33 | 7.74 ± 0.67 | 7.95 |
| B14 | Hybrid* | 8.56 ± 0.35 | 18.64 ± 0.35 | 8.71 ± 0.87 | 4.97 |
| B15 | Hybrid* | 9.49 ± 0.35 | 22.36 ± 0.36 | 7.59 ± 0.74 | 7.39 |
| B16 | B. tkazinoki | 7.44 ± 0.47 | 23.96 ± 0.34 | 7.95 ± 0.78 | 8.01 |
| B17 | Hybrid* | 8.21 ± 0.31 | 22.93 ± 0.36 | 8.68 ± 0.77 | 7.13 |
| B18 | Hybrid* | 8.53 ± 0.51 | 23.18 ± 0.34 | 7.98 ± 0.70 | 7.60 |
| B19 | B. kazinoki | - | - | - | - |
| B20 | Hybrid* | 9.53 ± 0.40 | 19.88 ± 0.52 | 7.40 ± 0.77 | 6.24 |
| B21 | Hybrid* | 9.40 ± 0.37 | 22.52 ± 0.63 | 8.26 ± 1.00 | 7.13 |
| B22 | Hybrid* | 11.17 ± 0.43 | 21.89 ± 0.68 | 8.65 ± 0.96 | 6.62 |
| B23 | Hybrid* | 9.15 ± 0.36 | 22.84 ± 0.69 | 9.40 ± 1.06 | 6.72 |
| B24 | Hybrid* | 7.60 ± 0.28 | 17.10 ± 0.57 | 5.58 ± 0.58 | 5.76 |
| Average | | 9.03 | 20.91 | 7.57 | 6.67 |
모든 시료의 섬유장, 섬유폭, 루멘폭, 세포벽 두께는 평균 9.03 mm, 20.91 μm, 7.57 μm, 6.67 μm로 확인되었다. 분자계통학적으로 닥나무와 꾸지나무의 교잡종으로 분류되더라도 섬유장이 최소 7.6 mm에서 최대 11.17 mm까지의 범위로 섬유장에 차이를 나타냈다. 섬유폭도 최소 17.10 μm에서 최대 23.64 μm까지로 비교적 넓은 범위의 길이와 폭이 확인되었다. 그러나 B. kazinoki인 B16시료는 섬유장이 7.44 mm, 섬유폭이 23.96 μm, 세포벽 두께가 8.01 μm로 모든 시료 중에서 가장 짧은 섬유장과 가장 두꺼운 섬유폭, 세포벽 두께를 나타내는 특징을 나타냈으며, 이러한 특징은 교잡종과 구분되는 특징이라고 판단된다.
섬유장은 종이의 지합, 인장 및 인열강도, 내절도 등에 큰 영향을 미치는 인자로 보고되었고 특히 섬유장이 증가함에 따라 인열강도와 내절도는 비례한다고 알려져 있다.13) 또한 세포벽의 두께가 얇으면 부피가 감소되고 투명하며 강도가 커지는 반면, 벽 두께가 두꺼우면 강도가 낮고 부피가 있으며 흡수도가 커진다고 보고하고 있다.23) 이러한 관계들로 교잡종인 닥나무로 종이를 제조하면 상대적으로 강도가 높은 종이가, B. kazinoki 닥나무로 종이를 제조하면 부피가 있으며 흡수도가 높은 종이가 제조됨을 유추할 수 있다. 이러한 특징을 토대로 모사나 회화 및 보존처리를 수행할 때 장인의 의도에 따라 종이를 선택할 수 있을 것으로 판단된다.
그리고 인피섬유의 형태학적 특징을 통해 펄프제지 특성을 runkel ratio(RR), slenderness ratio(SR), flexibility ratio(FR), rigidity coefficient(RC)를 적용하고 값을 도출하였다(Table5). 펄프제지 공정에서 RR, SR, FC, RC는 중요한 특성들로 여겨진다. 루멘폭과 세포벽의 두께로부터 기인하는 RR 값이 1 미만일 경우는 얇은 벽으로 간주되며 높은 기계적 강도를, SR 값은 내절도를 평가하는 지표로 사용된다. 섬유장과 비례 관계에 있는 SR 값은 0.7보다 낮으면 품질이 우수한 펄프, 종이 제조에 적합하지 않으며 제지용 섬유는 33 이상이 되어야 허용된다고 보고되고 있다. 그리고 FC와 RC 값은 강도 특성 중에서도 섬유결합의 정도와 상관관계가 있다. 상대적으로 높은 FC 값을 가지면 섬유 간 결합이 우수하여 비교적 인장강도와 파열강도, 내절도가 우수한 종이를 생산한다고 보고되고 있다. FC 값은 일반적으로 0.55-0.75의 값을 나타내며 값이 감소할수록 수피와 나무가 성숙하다고 알려져 있다. RC 값은 증가할수록 섬유결합이 감소된다고 보고된다.20-24)
Table 5.
Variation in the derived morphological ratios of the bast fibers according to the paper-mulberry
| Abbreviation | RR | SR | FC | RC |
|---|
| B01 | 2.19 | 460.63 | 0.31 | 0.69 |
| B02 | 1.87 | 430.96 | 0.35 | 0.65 |
| B03 | 1.65 | 486.16 | 0.38 | 0.62 |
| B05 | 1.77 | 430.44 | 0.36 | 0.64 |
| B06 | 1.67 | 490.54 | 0.37 | 0.63 |
| B07 | 1.95 | 477.45 | 0.34 | 0.66 |
| B08 | 1.84 | 491.20 | 0.35 | 0.65 |
| B09 | 0.93 | 490.54 | 0.52 | 0.48 |
| B10 | 2.22 | 418.54 | 0.31 | 0.69 |
| B11 | 2.13 | 375.50 | 0.32 | 0.68 |
| B12 | 2.14 | 455.97 | 0.32 | 0.68 |
| B13 | 2.05 | 395.52 | 0.33 | 0.67 |
| B14 | 1.14 | 459.23 | 0.47 | 0.53 |
| B15 | 1.95 | 424.42 | 0.34 | 0.66 |
| B16 | 2.01 | 310.52 | 0.33 | 0.67 |
| B17 | 1.64 | 358.05 | 0.38 | 0.62 |
| B18 | 1.90 | 367.99 | 0.34 | 0.66 |
| B19 | - | - | - | - |
| B20 | 1.69 | 479.38 | 0.37 | 0.63 |
| B21 | 1.73 | 417.41 | 0.37 | 0.63 |
| B22 | 1.53 | 510.28 | 0.40 | 0.60 |
| B23 | 1.43 | 400.61 | 0.41 | 0.59 |
| B24 | 2.06 | 444.44 | 0.33 | 0.67 |
| Average | 1.80 | 435.26 | 0.36 | 0.64 |
가장 얇은 세포벽을 갖는 B09 시료의 경우 RR값이 0.93으로 나타나 종이로 만들었을 때 가장 높은 기계적 강도를 가질 것으로 판단된다. 모든 시료의 SR 값은 33 이상으로 제지용 섬유로 적합하며, 분자계통학적 연구로부터 B. kazinoki로 판정된 B16시료는 가장 짧은 섬유장을 가지며, 이와 비례하는 관계인 SR 값도 310.52로 최저값을 갖는 특성을 보였다. 모든 시료의 FC 값은 0.31-0.52의 범위를 나타내었다. B09 시료의 FC 값이 0.52로 가장 큰 값을 가지며, 상대적으로 모든 시료 중에서 섬유가 손쉽게 접히며 표면 접촉 및 섬유와 섬유 간 결합이 우수할 것으로 판단된다. RC 값은 세포벽이 가장 얇은 B09 시료가 가장 낮았다. 또한 세포벽이 가장 두꺼운 B. kazinoki가 높은 RC 값을 가질 것으로 예상했으나, 섬유폭의 두께가 가장 길기 때문에 상쇄되어 0.67의 값을 나타냈다.
4. 결 론
이전 연구에서와 같이, 기존의 분류학 방법으로는 많은 오류가 존재함으로 이러한 문제점을 해결하기 위해서 본 연구에서는 전통한지 공방에서 재배되고 있는 전통한지의 주재료인 닥나무속 식물을 분자계통학적 분석을 통해 닥나무속 식물을 분류하고, 그에 따른 형태학적 특성을 얻었으며 결과를 요약하면 아래와 같다.
전통한지 공방에서 재배되고 있는 닥나무속 식물은 분자계통학적 분석을 통해 두가지 종류로 분류되었다. 첫번째로 채취한 대부분의 시료는 교잡종(B. kazinoki × B. papyrifera)으로 나타났다. 그리고 전체 24개체 시료 중 2개체만이 B. kazinoki로 분류되었다.
형태학적 특징으로, B16 시료(B. kazinoki)는 섬유장이 가장 짧고 가장 낮은 SR 값을 나타냈다. 이는 기계적 강도면에서 상대적으로 약한 성능을 가질 것으로 판단되지만, 섬유폭, 루멘폭, 세포벽에서 우수한 값을 가져 강도는 낮지만 부피감이 있고 흡수도가 높은 종이의 제조가 가능할 것으로 판단된다. 상대적으로 긴 섬유장을 가지는 교잡종은 좋은 기계적 강도를 나타낼 것으로 예측되며 특히 인열강도, 내절도에서 강도가 우수한 종이를 제조할 수 있을 것으로 판단된다.
한편, 본 연구에서 수행된 펄프·제지 특성은 단섬유에 해당하는 기준이며 장섬유인 닥나무 인피섬유를 가지고 전통방식으로 종이를 만드는 한지에 적합한 조건이라고 하기에는 어려움이 있다. 따라서 장섬유에도 적용할 수 있는 펄프·제지 특성 기준을 위한 연구가 앞으로 필요할 것으로 판단된다. 그리고 B. kazinoki인 닥나무의 수급 부족으로 단일시료가 사용되어 과학적 데이터의 신뢰도와 보편성 상승을 위한 추가연구가 요구된다.
그럼에도 불구하고 본 연구를 통한, 분자계통학적 분류에 따른 펄프·제지 특성이 다르다는 점은 전통한지를 제작할 때 닥나무를 선별하여 펄프·제지 특성을 고려한 전통종이의 제조가 가능할 것으로 보이며, 전통한지를 사용하고자 하는 사용자들의 목적과 용도에 맞게 전통종이의 기능과 특색을 통한 선택의 범위를 넓혀주는 기회가 될 것이다.
Acknowledgements
This study was conducted as part of the National Research Institute of Cultural Heritage R&D Project (Project number: NRICH-1905-A11F-1).
Literature Cited
Lee, Y. S., A study on the characteristics and suitability of broussonetia kazinoki for erosion control species [dissertation], Sangji University, pp.1-43 (2017).
Nakai, T., Flora Koreana Ⅱ, Imperial University of Tokyo. Tokyo, Japan. pp.193 (1911).
Mori, T., An enumeration of plants hitherto known from corea, 朝鮮總督府, pp.123 (1922).
Park, B. I., A study on the morphological characteristics of leaves of Korean mulberry trees, Jeonbuk National University Proceedings of Natural Sciences 11:13-14 (1969).
Ohba, H. and Akiyama, S., Broussonetia (Moraceae) in Japan, The Journal of Japanese botany 89:123-128 (2014).
Kim, M. Y., Kim, T. J. and Lee, S. T., A taxonomic study of the Korean Broussonetia (Moraceae) by multivariate analyses, Korean Journal of Plant Taxonomy 22(4):241-255 (1992).
10.11110/kjpt.1992.22.4.241Kim, H. C., Studies on the mechanical and functional properties of Hanji (Korean paper) yarn [dissertation], Jeonbuk National University, pp.1-124 (2007).
Go, I. H., Jo, A. H., Jeong, S. L., Heo, T. Y., Cho, K. and Choi, T.H., Predictive model of geographical origin discrimination of paper mulberry and handmade paper using ICP-AESMS and multivariate statistical analysis, Journal of Cultural Heritage 49:222-228 (2021).
10.1016/j.culher.2020.12.004Go, I. H., Jeong, S. H., Anatomical, morphological, and chemical characteristics of paper-mulberry wood and bast fiber for raw material of Korean paper (Hanji), Journal of Conservation Science 34(6):517-524 (2018).
10.12654/JCS.2018.34.6.07Seth, R. S, Fibre quality factors in papermaking - II The importance of fibre coarseness, materials interactions felevant to the pulp. Paper and Wood Industries, 197:1-143 (1990).
10.1557/PROC-197-143Lee, H. R., Paper science Gwangilmunhawsa, Seoul, pp.387-389 (1996).
Zhu, J. Y., Tim Scott, C., Scallon, K. L. and Myers, G. C., Effects of plantation density on wood density and anatomical properties of red pine (Pinus resinosa Ait.), Wood and Fiber Science 39(3):502-512 (2007).
Horn, R. A., 1978, Morphology of pulp fiber from hardwoods and influence on paper strength, Forest Service Research Paper, pp.1-12 (1978).
Allen, G. G., Flores-vergara, M. A., Krasynanski, S., Kumar, S. and Thompson, W. F., A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide, Nature Protocols, 1:2320-2325 (2006).
10.1038/nprot.2006.384Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S., Buxton, S., Cooper, A., Markowitz, S., Duran, C., Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data, Bioinformatics 28:1647-1649 (2012).
10.1093/bioinformatics/bts199Kimura, M., A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences, Journal Molecular Evolution 16(2):111-120 (1980).
10.1007/BF01731581Stamatakis A., RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with tho usands of taxa and mixed models, Bioinformatics 22:2688-2690 (2006).
10.1093/bioinformatics/btl446Ekhuemelo, D. O. and Udo, A. M., Investigation of variations in the fibre characteristics of Moringa Oleifera (Lam) stem for pulp and paper production, International Journal of Science and Technology 5(1):19-25 (2016).
Dutt, D. and Tydgi, C. H., Comparison of various eucalyptus species for their morphological, chemical, pulp and paper making characteristics, Indian Journal of Chemical Technology 18(2):145-151 (2011).
Ogbonnaya, C. I., Roy-Macauley, H., Nwalozie, M.C. and Annerose, D.J.M., Physical and histochemical properties of kenaf (Hibiscus cannabinus L.) grown under water deficit on a sandy soil, An International Journal of Industrial Crops and Products 7(1):9-18 (1997).
10.1016/S0926-6690(97)00034-4Choi, T. H., Jo, N. S., Lee, S. H. and Oh, S. G., Anatomical characteristics of mulberry bast fiber by location, Journal of Korea TAPPI 48(3):298-303 (2007).
Sadiku, N. A., Oluyege, A. O. and Ajayi, B., Fibre dimension and chemical characterisation of naturally grown Bambusa vulgaris for pulp and paper production, Journal of Bamboo and Rattan 15(1-4):33-43 (2016).


