

1. 서 론
범지구적인 화석에너지 사용 및 온실가스 배출의 증가에 따라 기후변화가 급격히 진행되면서 이를 해결하기 위하여 국가별 대체에너지 보급·확산 정책이 수립·시행되고 있다. 우리나라도 2020년부터 탄소 배출을 의무적으로 감축해야 하는 국가로 지정됨에 따라 에너지 공급량 대비 신재생에너지 비율을 2030년까지 11%로 확대하는 국가에너지기본계획을 수립하였다. 그러나 최근 국제에너지기구(IEA)의 발표에 따르면 우리나라는 OECD 회원국 중에서 1인당 탄소배출 증가율이 가장 높아 기후변화 대응 능력이 부족한 것으로 나타났다.1) 국가적 차원에서 의무 감축을 계획대로 이행하기 위한 신재생에너지 기술의 개발과 국내 산업에의 적용이 시급한 실정이다. 바이오연료 생산을 위한 2세대 바이오매스인 목질 바이오매스는 식량자원 기반의 1세대 바이오매스와 달리 윤리적 문제를 야기하지 않는 비식용 자원이고, 지구상 총 바이오매스 양의 90%를 차지할 만큼 풍부하고 재생가능한 바이오자원이라는 점에서 대체에너지원으로서의 잠재성이 높다. 우리나라는 국토의 약 64%가 산림으로 목질 바이오매스 자원이 풍부하지만, 직경 14 cm 이하인 소경목이나 해충 피해목은 목재 소재로의 활용이 제한된다. 이러한 미이용 목재를 원료로 하여 바이오연료를 생산한다면 폐기물 재활용 및 지속가능한 에너지 생산 모두를 충족시키는 자원이 될 것이다. 그러나 목질 바이오매스는 미세 구조에 있어 그 활용에 제약을 받는다. 목재의 세포벽은 D-글루코오스가 β-1,4-글리코시드 결합으로 이루어진 셀룰로오스, 주로 자일란 또는 글루코만난으로 구성된 헤미셀룰로오스, 그리고 분자구조가 복잡하여 세포벽을 단단히 유지하는 리그닌을 주성분으로 한다.2) 이들은 형태학적으로 매우 견고하게 결합되어 있어 화학물질의 접근이 어렵고, 셀룰로오스 분자 내 결정성 구조는 당화 저해인자로 작용할 수 있다.3) 따라서 이를 극복하기 위한 전처리 및 당화·발효 기술에 대한 연구가 꾸준히 진행되고 있다.
과초산은 산화력이 강한 화학물질로, 비교적 낮은 온도에서 리그닌 구조를 공격하여 목질 세포벽 구조를 파괴하는 전처리 방법으로 알려져 있다. 과초산에 의해 생성된 강한 친전자성을 띠는 옥소늄 이온(H3O+)이 리그닌 내 방향족 고리와 올레핀 측쇄 구조 등 전자가 풍부한 부분과 반응하여 전체적인 리그닌 구조가 파괴된다.4) 전처리 이후에는 목질 바이오매스의 화학적 및 구조적 특성을 고려한 당화를 실시해야 한다. 바이오에탄올 생산을 위한 당화 공정 중 효소를 이용하는 경우, 기질에 따라 cellulase와 hemicellulase의 종류가 다양하므로 목질계 셀룰로오스와 헤미셀룰로오스에 적합한 효소를 혼합하여 사용한다. 목질 바이오매스의 효소 당화는 endo- 및 exo- 형태의 ‘β-glucanase’와 셀로비오스를 선택적으로 분해하는 ‘β-glucosidase’가 서로 동시에 작용하여 목재 내 사슬 구조로 결합된 다당 구조를 단당 단위로 끊어내 글루코오스를 생산한다. 여기에 목재 내 헤미셀룰로오스의 구성 성분비에 따라 xylanase와 glucomannase를 적절히 배합하면 전체적인 효소 당화 효율을 높일 수 있다.5) 또한, 목질 바이오매스의 경우 수종별 구성 성분의 비율과 결합 양상이 다양하여 효율적 당화를 위해 투입해야 할 cellulase와 hemicellulase의 양이 다른데, 비교적 단가가 저렴한 hemicellulase를 조효소로 활용하여 효소 비용의 대부분을 차지하는 celllulase 양을 최소화하면서 전체적인 당화 공정에서의 비용을 줄일 수 있다.
본 연구에서는 과초산 전처리를 통해 리그닌 성분을 제거한 목질 바이오매스의 효소 당화 시 수종별 구성 성분 차이에 따른 공시 효소의 투입 최적 비율을 탐색하여 시료 특성에 따른 효소 당화 조건을 구명하고, 궁극적으로 목질 바이오매스의 기본 특성에 따른 최적 효소 당화 조건 탐색 시 활용할 수 있는 기초자료를 제시하고자 하였다.
2. 재료 및 방법
2.1 공시 재료
2.1.1 공시 목재
본 연구에서 사용한 목재 시료는 활엽수재 및 침엽수재 각 1종으로 국내 대표적 목재 생산 수종인 신갈나무(Quercus mongolica Fisch. ex Ledeb.)와 낙엽송(Larix kaempferi Lamb.)을 공시수종으로 하여 경기도 안양시에 위치한 서울대학교 농업생명과학대학 수목원에서 12년생 공시목을 선정, 벌채 후 목재 원판(wood disc)으로 제조하여 평형 함수율에 도달할 때까지 천연 건조하여 사용하였다.
공시목은 수피와 옹이 등 화학 조성이 달라질 수 있는 부분을 제거한 후 Wiley micro-mill(Thomas Scientific, USA)로 분쇄하여 40 mesh 이하의 목분을 선별하여 화학 조성분 분석 시료로 사용하였다. 1.0×2.0×0.5 cm3 크기의 목재 칩을 과초산 전처리 및 효소 당화 실험용 시료로 사용하였다. 공시목재의 항온항습 조건(23±1°C, RH 50±1%)에서의 평형 함수율은 신갈나무 및 낙엽송이 각각 8.53% 및 10.09%였다.
2.1.2 당화 효소
본 연구에서 목재의 당화 실험에 사용한 효소는 목질 바이오매스 당화용 상용 효소인 Novozymes CellicⓇ CTec2 및 HTec2로 각 효소의 사양 및 특성은 Table 1과 같다. 당화 실험에 앞서 CTec2 및 HTec2 내 cellulase 효소 활성도를 NREL 표준 시험방법(NREL/TP-510-42629)에 따라 FPU 단위로 측정하였으며,6) 각각 약 130 FPU/mL 및 75 FPU/mL로 측정되었다.
Table 1.
Characteristics of CellicⓇ CTec2 and HTec2 and their features
2.1.3 공시 시약
목질 바이오매스 시료 내 유기용매 추출성분 정량 분석에는 아세톤(CH3COCH3: 99.5%, Daejung, Korea)을 사용하였다. 산 불용성 및 가용성 리그닌 함량 분석 시 황산(H2SO4: J. T. Baker, USA)을 희석하여 사용하였으며, 중성 당 분석에서는 Bio-LC에 고정상으로 수산화나트륨(50%(w/w) NaOH solution, Fisher Chemical, USA)을 희석하여 적용하였다. 과초산 전처리에는 아세트산(CH3COOH: J. T. Baker, USA)과 과산화수소(30% H2O2 solution, Junsei Chemical, Japan)를 혼합하여 사용하였다. 효소 당화 시 효소 활성 pH를 맞추기 위해 구연산나트륨(Na3C6H5O7·2H2O: Duksan, Korea)과 구연산(C6H8O7: Duksan, Korea)을 혼합한 구연산나트륨 완충용액(1 M, pH 5.5)을 사용하였고, 방부제로 아지드화 나트륨(NaN3: Daejung, Korea)을 희석하여 사용하였다.
2.2 실험 방법
2.2.1 화학 조성분 분석
2.2.1.1 추출성분 함량
공시 목재의 세포벽을 구성하는 성분 중 유기용매 추출성분의 총 함량을 분석하기 위한 실험은 TAPPI T 204 cm-97에 의거하여 실시하였으며, 추출 용매로 아세톤을 사용하였다. 추출 후 탈지된 시료와 원통여과지는 105±1°C에서 24시간 이상 건조시킨 후 전건 중량을 측정하였다. 추출액이 담긴 둥근 플라스크는 회전식 농축기(HS-2000N, Hahn Shin Scientifics, Korea)를 이용하여 아세톤을 회수한 후, 잔여 아세톤은 후드 내에서 건조하였다. 둥근 플라스크는 105±1°C에서 24시간 이상 건조 후 전건 중량을 측정하였다.
2.2.1.2 리그닌 함량
공시 목재 내 리그닌 함량 측정은 TAPPI T 222 om-02에 의거하여 실시하였다. 72%(w/w) 황산으로 가수분해한 산물을 이용하여 산 불용성 리그닌 및 산 가용성 리그닌의 함량을 구하고 그 합을 리그닌의 총 함량으로 계산하였다. 가수분해 산물을 1차 여과한 여액을 UV/Vis spectrometer(Lambda 35, PerkinElmer, USA)를 이용하여 205 nm에서의 흡광도(0.2-0.7)를 측정하여 TAPPI UM 250의 계산식에 따라 산 가용성 리그닌을 정량 분석하였으며, 여과 산물은 105±1°C에서 24시간 이상 건조 후 전건 중량을 측정하여 산 불용성 리그닌을 정량하였다.
2.2.1.3 중성 당 조성 및 함량
공시 목재 내 중성 당(아라비노오스, 갈락토오스, 글루코오스, 자일로오스, 만노오스) 조성 및 함량 분석은 NREL 표준 시험방법(NREL/TP-510-42618)에 의거하여 실시하였으며, 산 가수분해 여액을 HPLC(ICS-3000, Dionex, USA)를 이용하여 분석하였다. HPLC 분석 조건은 Table 2와 같다.
2.2.2 과초산 전처리
공시 목재의 탈 리그닌 처리 및 효소 접근성 향상을 목적으로 과초산 전처리를 실시하였다. 예비실험 결과를 바탕으로 탈리그닌율이 거의 100% 이루어지면서 시료 내 글루코오스 손실이 가장 적은 다음의 조건으로 전처리를 실시하였다.
전건 기준 약 30 g의 목재 칩이 들어있는 1 L 유리병에 전처리 약액(CH3COOH:30% H2O2=3:7(v/v) 혼합액) 300 mL를 투입하고, 진탕 항온수조(DS-250SW, Daewon Science Co. Ltd., Korea)를 이용하여 70±1°C, 120 rpm 조건에서 6시간 동안 전처리를 진행하였다. 전처리 후 시료는 증류수로 충분히 수세하여 중화시키고, 해리기를 이용해 목질 섬유소를 해리하여 수집한 후 함수율과 전수율을 측정하였다.
2.2.3 효소 당화
과초산 전처리를 통해 얻은 목질 섬유소를 기질을 대상으로 효소 당화 시험을 실시하였다. 100 mL 삼각플라스크에 전처리가 완료된 목질 섬유소 전건 중량 약 1 g을 투입하고, 구연산 나트륨 완충용액(1 M, pH 5.5) 5 mL, 아지드화 나트륨 수용액(1%, v/v) 1.3 mL를 가했다. 이후 효소 칵테일을 포함하여 총 중량이 40 g이 되도록 하였다. 실험 목적에 따라 효소 칵테일의 양은 다르게 투입하였으며, 해당 조건은 Table 3과 같다. 효소 당화는 진탕 배양기(HB-201SI, Hanbaek Science Co. Ltd, Korea)에서 50±2°C, 150 rpm의 조건으로 회전식 진탕배양을 통해 실시하였으며, 각 반응기로부터 3, 6, 9, 12, 24, 48, 72시간의 반응시간별 배양액을 마이크로튜브에 약 0.5 mL씩 채취하였다. 채취한 배양액이 담긴 마이크로튜브를 즉시 105°C로 가열된 실험용 가열반응기(DMB-3, Misung Scientific Co. Ltd., Korea)에 옮겨 5분간 효소 가수분해 반응을 정지시킨 후, 5분간 10,000 rpm 조건으로 원심분리기(Smart 15, Hanil Science, Korea)에서 분리 후 상등액을 취해 당 분석 시료로 사용하였다.
Table 3.
Various enzyme ratio and dosages in the enzymatic saccharification
2.2.4 효소 당화 잔사 분석
당화 과정에서 시료 내 탄수화물 성분과 당화 양상의 관계를 살펴보기 위해 당화 잔사를 산 가수분해하여 반응 시간별 당화 잔사 내 당 조성 변화를 분석하였다. 예비 실험을 통해 1 g 상당량의 시료 내 글루코오스를 최대로 당화시킬 수 있는 효소 투입량을 고정하고, 반응 시간별 당화 잔사 내 당 함량 변화를 파악하여 그 영향을 비교 분석하였다. 수종별 최종적으로 결정된 혼합비의 효소를 가한 반응기를 반응시간별로 준비하고, 미리 전건 중량을 기록한 여과지(Whatman No. 1)를 활용해 감압 여과하였다. 여과된 당화 잔사는 증류수를 활용해 충분히 수세 후, 전술한 바와 같이 산 가수분해하여 중성 당 분석을 실시하였다.
3. 결과 및 고찰
3.1 과초산 전처리 목질 바이오매스의 특성
바이오에탄올 생산에 있어 목질 바이오매스는 수종별로 다른 화학 조성에 따라 전처리 및 당화 효율이 다르게 나타난다. 따라서 우선적으로 전처리 전후의 화학 조성 변화 분석을 통해 과초산 전처리가 목질 바이오매스 화학 조성에 미치는 영향을 분석하고, 효소 당화 기질이 되는 전처리 후 목질 섬유소의 화학 조성 변화를 관찰하였다. 목질 바이오매스 내 셀룰로오스와 헤미셀룰로오스는 산 가수분해 시 단당으로 분리될 수 있으며, 이를 정량 분석하면 각 조성분의 비율을 구할 수 있다. 과초산 전처리를 실시하기 전 공시 목재의 기본 화학 조성 분석 결과는 Table 4와 같다. 이 때 산 가용성 리그닌의 양은 전체 구성성분의 1% 이하의 미량으로 확인되어 산 불용성 리그닌의 양과 함께 나타냈다. 신갈나무 및 낙엽송 목재 칩의 과초산 전처리 후 목질 섬유소의 전수율은 각각 60.4% 및 54.8%로 침엽수인 낙엽송에서 더 많은 바이오매스 감소율을 나타냈으며, 중성 당 및 리그닌 정량 분석 결과 목질 세포벽 내 리그닌 및 헤미셀룰로오스가 전처리 과정에서 우선적으로 분해되어 발생한 차이로 사료된다. 리그닌은 세포벽의 외벽에 주로 분포하여 과초산에 의해 가장 먼저 분해되는 주성분으로,7) 본 실험에서도 과초산 전처리 후 두 수종 모두에서 리그닌이 대부분 분해된 것을 확인할 수 있었다. 이 후 중성 당 분석 시 헤미셀룰로오스 유래 단당(활엽수의 경우 자일로오스, 낙엽송의 경우 아라비노오스, 갈락토오스, 자일로오스, 만노오스)이 낙엽송에서 상대적으로 더 많이 손실된 결과로 미루어 볼 때(Fig. 1), 탄수화물 성분 중 셀룰로오스보다 헤미셀룰로오스 성분이 과초산에 의해 먼저 가수분해 되었고 이는 낙엽송에서 더욱 활발하게 이루어졌음을 확인할 수 있었다.
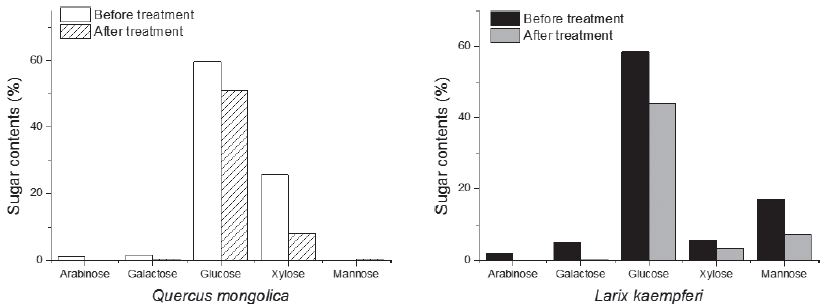
3.2 과초산 전처리 목질 바이오매스의 효소 당화 특성
3.2.1 효소별 당화 특성
CTec2 및 HTec2의 효소 당화 특성을 파악하기 위해 각 효소를 단독 처리하여 당화율 차이를 비교하였고, 그 결과 당화액에서 글루코오스와 자일로오스가 주로 검출되었다(Figs. 2와 3). 이 때 낙엽송 목재 칩의 화학 조성 분석 시 자일로오스보다 많았던 만노오스 성분이 당화액 내 존재하지 않는 것으로 나타났다. 이를 통해 CTec2 및 HTec2 내 O-acetyl-galactoglucomannan을 단당으로 분해하는 효소가 부재함을 알 수 있었고, 이 때문에 분해되지 않은 헤미셀룰로오스가 효소 반응을 저해할 가능성을 고려할 수 있었다. CTec2 단독 처리 시 신갈나무 목채 칩에서의 최종 글루코오스 전환율은 낙엽송보다 약 5% 정도 높게 나타났다. 이는 전처리 후 잔존 헤미셀룰로오스의 양이 상대적으로 많은 낙엽송에서 cellulase가 가수분해 반응에 대한 저해 영향을 받기 때문인 것으로 사료된다. 또한, 당화가 진행되는 동안 낙엽송 섬유소의 가수분해 속도가 신갈나무 섬유소에 비해 느린 것을 볼 때, 헤미셀룰로오스 내에서 당화 효소에 의해 분해되지 않는 만난(mannan) 성분에 의해 효소 반응이 저해받는 것으로 사료되었다.8) Cellulase가 상대적으로 적게 함유된 HTec2를 사용했을 때에는 두 수종의 섬유소 간 글루코오스 전환율이 7%까지 차이가 발생하는 것을 볼 때, cellulase 함량이 적을수록 저해 영향이 커지는 것으로 사료되었다. 또한, CTec2의 경우 HTec2와 달리 24시간 이내에 글루코오스 전환이 완료되는 결과를 나타냈는데, 이는 CTec2 내에 존재하는 β-glucosidase가 셀로비오스를 분해하면서 저해인자의 감소로 발생하는 효소 당화 속도 차이에 기인한 것으로 사료된다.
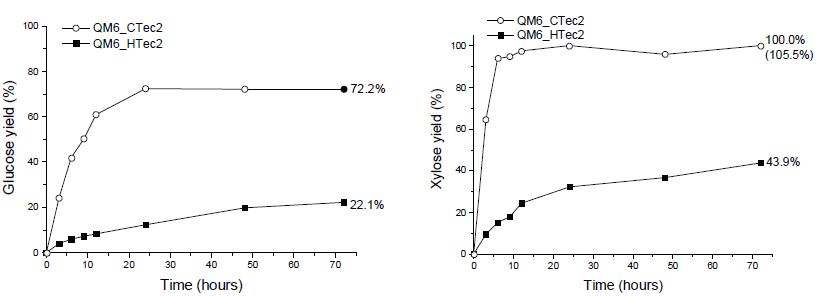
Fig. 2.
Changes in glucose and xylose yields during enzymatic saccharification of pretreated Quercus mongolica fibers with CTec2 and HTec2.
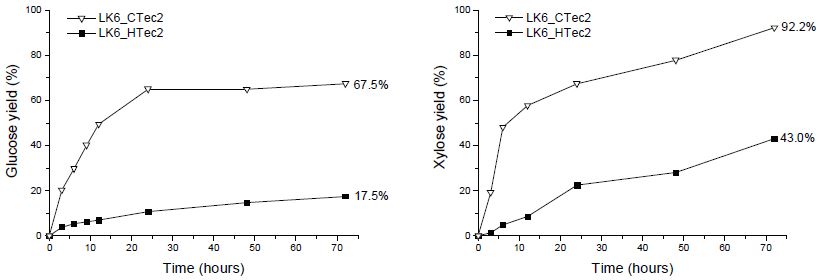
Fig. 3.
Changes in glucose and xylose yields during enzymatic saccharification of pretreated Larix kaempferi fibers with CTec2 and HTec2.
자일로오스의 경우, 신갈나무 섬유소와 낙엽송 섬유소에서 당화 양상에 차이를 보였다. 신갈나무 섬유소에서는 투입된 CTec2 내 존재하는 hemicellulase만으로도 24시간 내 자일로오스가 100% 가까이 전환되는 것으로 나타났다. 낙엽송 섬유소에서는 원 시료 내 자일로오스가 6%임에도 불구하고 효소의 당 전환 속도가 신갈나무에 비해 더딘 것으로 나타났으며, 이는 낙엽송 내 헤미셀룰로오스 성분을 이루는 O-acetyl-galactoglucoman-nan을 분해하는 효소가 당화액 내 잔여 효소에 의해 가수분해 반응이 저해되기 때문으로 사료된다.
3.2.2 효소 투입량에 따른 당화 특성
효소는 투입량이 늘어날수록 가수분해 수율이 증가하나, 과량 투입 시 바이오연료 생산 공정에 필요한 당화효소 비용의 부담이 커지고 효소 당화 과정에서 불필요한 저해요소가 발생할 수 있다. 따라서 CTec2 투입량에 따른 글루코오스 수율 등 효소 당화 특성을 관찰, 최적 투입량을 알아보고자 하였다. 두 수종에서 CTec2의 최적 투입량은 다르게 나타났다(Fig. 4). 신갈나무의 경우 0.14-0.15 mL(18.2-19.5 FPU) 투입 시 최고 글루코오스 수율을 나타냈으며, 낙엽송에서는 0.16-0.17 mL(20.8-22.1 FPU) 투입 시 가장 높은 글루코오스 수율을 나타냈다. 신갈나무의 경우 72시간 내 최고 당 수율(96.2%)이 낙엽송의 최고 당 수율(90.9%)보다 높았으며, 12-24시간 사이에 이미 대부분의 가수분해 반응이 종결되는 양상을 확인할 수 있었다. 이는 화학 조성분 함량 차이로 인한 것으로 설명할 수 있으며, 선행 연구보고9)에서 증명한 침엽수와 활엽수 간 공극 용적 차이로 인한 효소 접근성 차이 또한 이에 영향하는 것으로 사료되었다.
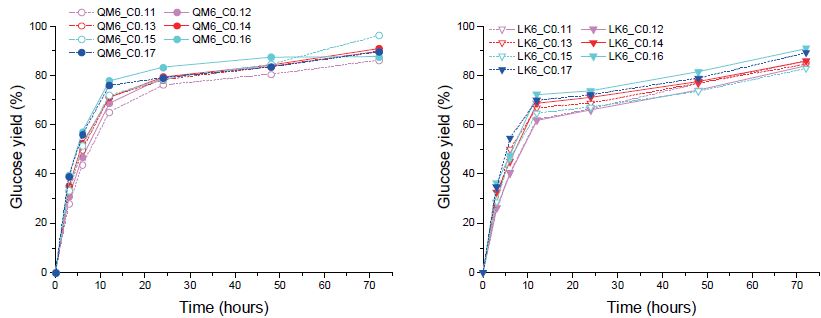
3.2.3 효소 혼합 비율에 따른 당화 특성
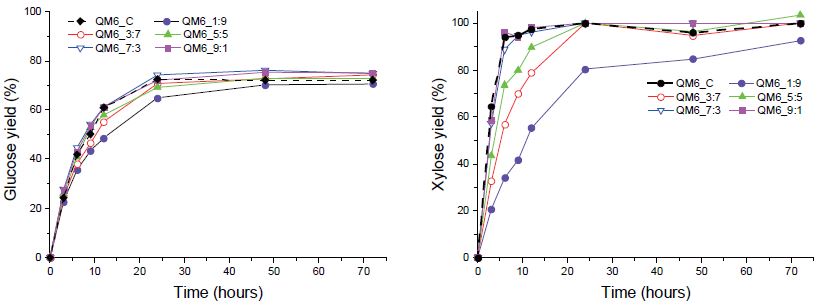
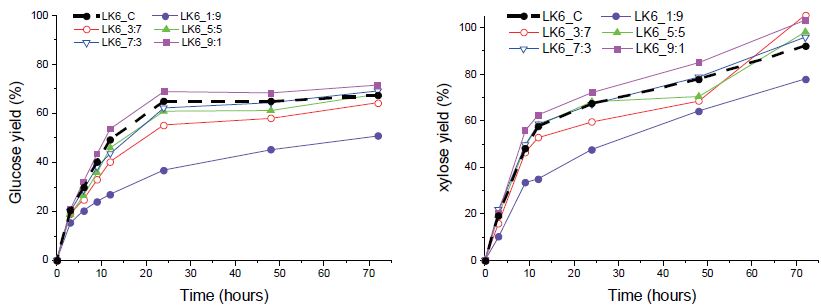
목질 바이오매스 내 구성성분이 다양함에 따라 효소 당화 시 반응하는 기질은 복합적으로 작용할 수 있다. 또한, 서로 다른 두 효소를 혼합하여 투입하였을 때 가수분해 반응의 속도나 최종 당 수율이 두 효소의 가수분해 특성과 다를 수 있으며, 이는 ‘복합효소 속도론’으로 밝혀진 바 있다. CTec2와 HTec2의 혼합 시 발생하는 가수분해 특성과 각각의 단독 투입 시 특성을 비교하기 위해 효소 투입 비율(CTec2:HTec2)을 1:9, 3:7, 5:5, 7:3, 9:1로 각각 다르게 하여 효소 당화를 실시하고, CTec2 단독 처리 시의 효과와 비교 분석하였다.
신갈나무에서는 HTec2를 일정량 이상 투입 시 CTec2보다 더 높은 당 전환율을 보이는 것으로 나타났다(Fig. 5). CTec2와 HTec2를 3:7, 5:5, 7:3 비율로 혼합한 효소 칵테일 활용 시 CTec2만 사용했을 때보다 최종 글루코오스 전환율이 0.4-2.6% 가량 높아졌으며, 특히 7:3으로 혼합 시에는 글루코오스 전환 속도 또한 빨라지는 것으로 나타났다. 자일로오스의 경우 CTec2 투입량에 비례하여 당화 속도가 빨라지는 경향을 나타냈다. 이로써 HTec2 투입 시 조효소 내 endoxylanase가 CTec2 내 xylanase와 함께 헤미셀룰로오스에 작용, 당화 상승효과를 나타내 헤미셀룰로오스 분해 효소 활성을 돕는 것을 확인할 수 있었다. 한편 낙엽송에서는 신갈나무와 다른 효소 당화 경향을 나타냈다(Fig. 6). HTec2 혼합 사용 시 CTec2의 글루코오스 수율이 높아졌으나, CTec2의 비율이 적어질수록 글루코오스 수율 또한 감소하는 것으로 나타났다(71.6%>69.2%>67.9%>64.4%>50.9%). 자일로오스 전환율 또한 혼합 후 CTec2 양에 비례하여 증가하는 경향을 나타냈다. 상기 실험 결과를 종합하면 낙엽송에서는 자일란의 함량이 적어 HTec2의 조효소로서의 효과가 미미하고 주성분의 분해가 CTec2에 의해 주도되지만, 신갈나무에서는 일정량의 HTec2 혼합 시 당 수율 또는 반응 속도를 높여 당화 효율을 상대적으로 증대시킬 수 있을 것으로 사료되었다.
3.3 Cellulase 활성도 및 혼합 효소에 따른 당화 특성
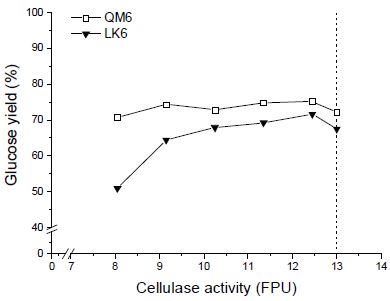
효소 당화 시 서로 다른 두 효소를 혼합하여 투입하였을 때 해당 반응의 속도나 최종 당 수율이 두 효소의 분해능의 합과 다를 수 있는데, 이 현상을 효소 간 ‘상승작용’이라고 한다. CTec2와 HTec2의 혼합 시 발생하는 상승작용을 확인하기 위해 앞서 효소 투입 비율을 달리한 효소 당화 실험에서의 글루코오스 수율과 각 혼합 효소의 cellulase 활성도(FPU) 합과의 관계를 확인하였다. 측정값을 바탕으로 cellulase 활성도와 글루코오스 수율의 관계를 살펴본 결과, 수종에 따라 두 측정값의 연관성이 다르게 나타났다(Fig. 7). 두 수종 모두에서 CTec2 단독 투입 시보다 HTec2와 혼합 투입 시 적은 FPU에서도 더 높은 당 수율을 나타내는 것으로 확인되었다. 실제 cellulase의 활성도와 글루코오스 수율은 비례하는 것이 일반적인데, 낙엽송의 경우 HTec2가 투입되더라도 CTec2 단독 투입 시와 동일한 양상을 보였으나 신갈나무에서는 cellulase 활성도 값에 큰 영향을 받지 않고 당화율이 최적 당화율과 비슷한 수준으로 유지되는 모습을 나타냈다. 이는 헤미셀룰로오스가 대부분 자일란으로 이루어져 있는 신갈나무 섬유소에서 HTec2 내 endoxylanase에 의해 셀룰로오스의 가수분해가 높은 상승효과를 얻는 것으로 사료되었다. 이러한 결과는 향후 cellulase 투입량을 줄이고도 최고 당 수율에 도달하는 경제적 효소 당화 공정 조건을 수립할 수 있는 가능성을 확인한 것으로 해석할 수 있다.
3.4 수종별 화학 조성 차이에 따른 효소 당화 특성
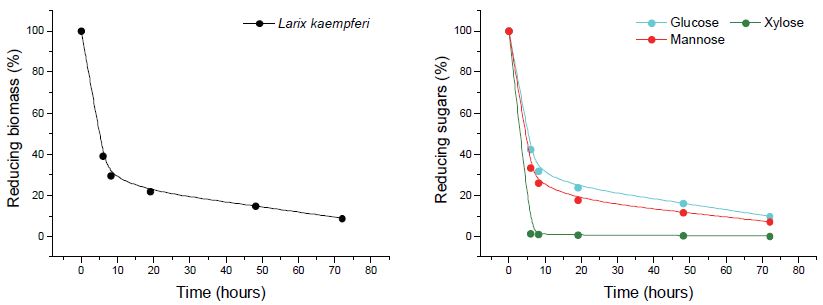
수종별 효소 당화 양상이 다르게 나타남에 따라 두 수종의 화학적 조성분과 효소 당화 특성간 관계를 파악하고자 하였다. 특히 두 수종 중 만노오스 함량이 상대적으로 높은 낙엽송에서 HTec2의 상승효과가 가장 미미한 것으로 나타났기 때문에 효소 당화에 미치는 만난계 물질의 영향을 살펴보기 위해 반응시간별 낙엽송의 당화 잔사 내 단당 함량의 변화를 살펴보았다(Fig. 8). 효소 가수분해 반응시간에 따른 낙엽송의 당화 잔사 내 중성 당 성분의 분해는 모든 당 성분을 대상으로 이루어졌다. 특히 효소 접근성 차이에 의해 셀룰로오스보다 헤미셀룰로오스에서 효소 가수분해가 진행됨에 따라 자일란계 탄수화물은 12시간에 기존 함량의 1% 미만까지 분해되는 것을 확인할 수 있었다. 그러나 만난 성분은 헤미셀룰로오스의 구성 성분임에도 불구하고 72시간에 거쳐 꾸준히 분해되는 것으로 나타났다. 또한, 낙엽송의 galactoglucomannan을 포함한 만난계 탄수화물은 효소에 의해 단당 형태로 분해되지 않지만, 효소에 의한 다른 구성 성분의 분해로 구조가 와해되어 당화액 내에 다당 및 이당류 형태로 존재하고 있음을 유추할 수 있었다. 또한, 글루코오스와 만노오스의 감소 양상이 비슷한 것으로 볼 때 글루코오스는 만난의 당화에 영향을 받고 있는 것으로 사료되었다.
4. 결 론
본 연구에서는 이용률이 저조한 국산 소경재의 바이오에탄올 생산 활용을 위해 활엽수인 신갈나무와 침엽수인 낙엽송 목재 칩을 대상으로 강력한 산화능을 가진 과초산 전처리를 시행하였다. 과초산 전처리에 의해 제조한 목질 섬유소를 대상으로 상용 당화 효소를 이용하여 효소 가수분해를 실시, 수종별 효소 당화 특성을 비교 분석하여 수종의 화학적 조성 및 특성에 따라 효율적인 글루코오스 당화 조건에 영향을 주는 인자를 구명하고자 하였다.
과초산 전처리 시료에 대한 CTec2 및 HTec2의 효소 당화 특성을 파악하기 위해 단독 투입 시 당화율 차이를 비교할 때, CTec2를 동일 양 투입 시 글루코오스 및 자일로오스 수율에서 신갈나무가 낙엽송보다 높은 것으로 나타났다. 특히 자일로오스 수율에서는 CTec2만으로도 신갈나무에서 12시간 이내에 시료 내 거의 대부분의 자일란이 분해되는 양상을 나타냈다. 또한, HTec2 단독 투입 시 실험 결과를 통해 조효소인 HTec2는 자일로오스의 분해 속도를 증진시키는 것으로 확인되었다. CTec2 단독 투입으로 최대 당 수율을 얻고자 했을 때 글루코오스가 많이 남아있는 신갈나무에 비해 낙엽송이 더 많은 효소를 필요로 하였으며, 이는 시료 내 헤미셀룰로오스 성분이 더 많이 잔존해있기 때문으로 사료되었다. 두 효소를 혼합하여 당화를 실시한 결과를 종합하면, 목질 바이오매스를 주 당화 대상으로 제조한 CTec2는 바이오매스 내 자일란의 함량에 따라 조효소를 활용하여 그 투입량을 줄일 수 있었다. 즉, CTec2에 endoxylanase를 함유한 HTec2를 투입함으로써 목질 바이오매스의 당화 속도 및 최종 당화율을 상승시켜 당화 효율을 높일 수 있으며, 전체 공정의 비용을 결정하는 CTec2의 투입량을 줄여 경제성을 확보할 수 있을 것으로 사료되었다. 이러한 효소 당화 특성 차이의 원인을 구명하기 위해 효소 반응시간에 따른 당화액 및 당화 잔사의 화학적 성분 분석을 실시하였다. 그 결과 글루칸이 분해되면서 다당인 만난계 물질도 간접적으로 저분자화되어 다당 또는 이당류로 당화액 내에 용해되는 것을 확인할 수 있었고, 이 물질들이 cellulase의 활성에 부정적인 영향을 주어 두 수종에서 글루코오스 당화 양상이 다른 것으로 결론지을 수 있었다.
모든 실험 결과를 종합하였을 때, CTec2와 HTec2를 활용한 목질 섬유소의 효소 가수분해 시 CTec2는 단독적으로 목질 섬유소를 가수분해할 수 있는 능력이 있다. 그러나 CTec2와 함께 HTec2를 조효소로 사용하였을 때 당화 시간을 단축시킬 수 있고, 단독으로 효소 가수분해했을 때보다 높은 당 수율을 기대할 수 있는 것으로 확인되었다. 이러한 두 효소의 상승효과는 헤미셀룰로오스 성분 중 자일란이 많은 수종일수록 더 뚜렷하게 나타나며, 만난이 많을 때에는 만노비오스, 만노오스 등에 의해 글루코오스 당 수율이 오히려 감소하는 양상을 보이므로 목질 섬유소의 화학 조성에 따라 HTec2의 투입량을 결정해야 한다.
